
Significado del nombre: El Tyrannosaurus Rex fue nombrado por Henry
Fairfield Osborn en 1905 y su nombre
significa “Rey de los Lagartos Tiranos", viene del griego tyrannus
“tirano” y saurus “lagarto”, y el latín rex “rey”, hace referencia a que era el
mayor dinosaurio carnívoro conocido cuando fue descubierto.
Clasificación científica:
Reino: Animalia
Filo: Chordata
Clase: Sauropsida
Subclase: Diapsida
Superorden: Dinosauria
Orden: Saurischia
Suborden: Theropoda
Superfamilia: Tyrannosauroidea
Familia: Tyrannosauridae
Subfamilia: Tyrannosaurinae
Género: Tyrannosaurus
Especies:
Tyrannosaurus Rex
Período que hábito: Maastrichtiense, Cretácico Superior,
aproximadamente hace 70,6 a 66,043 millones de años.
Localización actual: Estados Unidos (Colorado, Montana, Nuevo
Mexico, Dakota del Norte, Dakota del Sur, Texas y Wyoming) y Canadá (Alberta y
Saskatchewan).
Hábitat: habitaba en planicies de inundación y bosques
subtropicales húmedos donde acechaba a sus presas, en zonas demarcadas por
ríos, lagos y bosques exuberantes llenos de cicadáceas, helechos, plantas
florecidas y árboles como las coníferas, sicomoros y araucarias.
Dimensión: el mayor espécimen cuasicompleto, FMNH PR2081, apodado
Sue, midió 12,3 metros de largo, y 4 de alto hasta las caderas, el mayor cráneo
de Tyrannosaurus encontrado mide 1,5 metros de largo, más grande que el cráneo
Sue que mide 1,41 metros de largo lo que sugieren un tamaño potencial superior
de alrededor de 13 metros de largo.
Tipo de alimentación: Carnívoro, posiblemente hadrosáuridos y ceratópsidos,
aunque algunos expertos han sugerido que era principalmente carroñero.
Tyrannosaurus rex es la única
especie de Tyrannosaurus, un género monotípico de dinosaurio terópodo
tiranosáurido. Vivió en lo que es hoy Norteamérica occidental, con una
distribución mucho más amplia que otros tiranosáuridos. Comúnmente abreviado
como T. rex, fue uno de los últimos dinosaurios en existir antes de la
extinción masiva del Cretácico Terciario.

Tamaño
Aunque otros terópodos rivalizan
o superan al Tyrannosaurus rex en tamaño, todavía es el mayor tiranosáurido
conocido y uno de los mayores depredadores conocidos de la Tierra, midiendo
hasta 12,3 metros de largo, 4 metros de altura hasta las caderas y con un peso
estimado entre 6 a 9 toneladas, Tyrannosaurus rex era más largo que el bien
conocido terópodo del Jurásico Allosaurus y que el Carcharodontosaurus
africano, era ligeramente más pequeño que otros terópodos del Cretácico como
Spinosaurus y Giganotosaurus.

Morfologia

Como otros tiranosáuridos, T. rex
fue un carnívoro bípedo con un enorme cráneo equilibrado por una larga y pesada
cola, las vértebras se extienden desde la base del cráneo hasta la punta de la
cola, cada vértebra tenía un propósito específico.
El cuello del Tyrannosaurus formaba
una curva natural con forma de S como en otros terópodos, era grueso, musculoso
y corto para soportar su enorme cabeza, las vértebras del cuello ubicadas desde
la base del cráneo hasta la parte superior de la caja torácica, apoyan el
movimiento de lado a lado y de arriba a abajo de la cabeza del dinosaurio. Las
espinas en la parte superior de las vértebras añaden apoyo a los músculos que
controlan el movimiento de la cabeza. Las vertebras del cuello tienen forma de hendidura
en la parte inferior, dan mayor fuerza a las inserciones musculares, así como
la protección de los vasos sanguíneos vitales. La médula espinal pasa a través
de las vértebras del cuello y en el resto del cuerpo. Lugares ásperos en los
huesos ayudan a los paleontólogos a determinar dónde se adjuntan los músculos.

Las vértebras de las costillas o columna
vertebral se extendían desde la base del cuello hasta la cadera. Las espinas de
estas vértebras son más altas y más fuertes que las vertebras del cuello lo que
permite una mayor inserción muscular, asi mismo, la cara de la parte posterior de estas son mas
fuertes para soportar el enorme peso de la masa corporal de los dinosaurios.
Las costillas protegen los órganos vitales del dinosaurio, como el corazón, los
pulmones y el tracto digestivo.

El tiranosaurio poseía un gran
cráneo de 1,50 m aproximadamente, provisto de fenestras (aberturas) oculares y
nasales, reducían el peso y proporcionaban lugares para la inserción muscular,
como se ve en todos los terópodos carnívoros.

En otros aspectos el cráneo de
Tyrannosaurus era perceptiblemente diferente de los terópodos no tiranosáuridos
grandes, era extremadamente ancho en la parte posterior pero tenía un hocico
estrecho, permitiendo una visión binocular inusualmente buena.

Detalle de la visión
binocular del T.rex
El cráneo presenta un gran número
de huesos fusionados, supliendo la movilidad por una estructura más maciza,
cosa inusual en los terópodos, que por lo general tenían huesos ligeros, muchos
de estos huesos estaban neumatizados, conteniendo un panal de abejas de
espacios aéreos minúsculos que pueden haber hecho los huesos más flexibles
además de aligerarlos.

Éstas y otras características de
consolidación del cráneo son parte de la tendencia de los tiranosáuridos hacia
una mordedura cada vez mayor, que sobrepasaba fácilmente a todos los no
tiranosáuridos. La mandíbula que era capaz de abrir muy ampliamente tenia el
extremo de la quijada superior tenía forma de U, mientras que en la mayoría de
los carnívoros no tiranosáuridos las quijadas superiores tenían forma de V, lo
que aumentaba la cantidad de tejido y hueso que un tiranosaurio podría arrancar
con una mordedura, aunque también aumentara las tensiones en los dientes
delanteros.
Se ha afirmado que este
dinosaurio tuvo una de las mordeduras más poderosa de todos los depredadores,
con una fuerza de presión de más de 4 toneladas.
Con mas de 60 dientes, el
Tyrannosaurus rex muestran una marcada heterodoncia (diferentes en forma). Los
dientes premaxilares al frente de la mandíbula superior estaban estrechamente
empaquetados, con una sección de corte en forma de D, tenían refuerzo de cantos
en la superficie posterior y eran incisiformes (tenían láminas como cinceles) y
curvados hacia atrás. La sección de corte en forma de D, los bordes reforzados
y la curvatura hacia atrás reducían el riesgo que los dientes se rompieran
cuando el Tyrannosaurus mordiera y desgarrara. El resto de los dientes eran
robustos, conicos similares a dagas, estaban más espaciados entre sí y también
tenían refuerzo en los bordes. Los de la quijada superior eran más grandes que
los de la parte trasera de la quijada inferior.
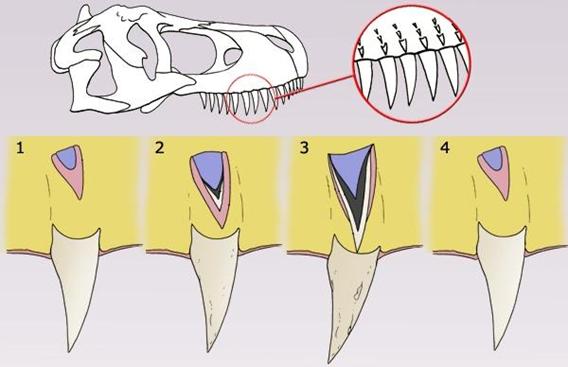
Proceso de reemplazo
de los dientes del T.rex
El mayor diente encontrado hasta
ahora se estima que medía 30 centímetros de largo incluyendo la raíz cuando el
animal estaba vivo, haciendo de este diente el mayor de cualquier dinosaurio
carnívoro descubierto hasta la fecha.

Los miembros superiores del
Tyrannosaurus eran pequeños, pero inusualmente fuertes para su tamaño y
terminaban en dos dedos con garras, junto con un pequeño metacarpiano
adicional, vestigio de un tercer dígito, la función de las extremidades
superiores es objeto de discusión. En 1906 Osborn especuló que podrían haber
servido para atrapar a la pareja durante la copulación. También se ha sugerido
que los miembros anteriores se utilizaron para ayudar a los animales a
levantarse de una posición de decúbito esternal.

Otra posibilidad es que los
miembros anteriores sujetaran a la presa durante la lucha, mientras las enormes
mandíbulas del tiranosaurio la mataban. Esta hipótesis se apoya en el análisis
biomecánico. Los huesos de las extremidades anteriores de Tyrannosaurus rex
presentan un hueso cortical muy grueso, que indica que se desarrollaron para
soportar cargas pesadas. El músculo bíceps braquial de un Tyrannosaurus rex
adulto era capaz de levantar 199 kg por sí solo, número que aumentaría al
actuar con otros músculos, como el músculo braquial. Por otra parte, el
antebrazo de Tyrannosaurus tenía una libertad de rotación limitada, con el
hombro y codo permitiendo giros de sólo hasta 40 y 45 grados, respectivamente.
En comparación, las mismas dos articulaciones en el Deinonychus permitían
movimientos de hasta 88 y 130 grados, respectivamente, mientras que un brazo
humano puede girar 360 grados en el hombro y moverse alrededor de 165 grados a
nivel del codo. Los pesados huesos de las patas delanteras, la fuerza extrema
de sus músculos y la rotación limitada pueden indicar un sistema evolucionado
para mantenerse firme a pesar de las tensiones provocadas por una presa que
forcejea por liberarse.

Rango de movimiento de la mano del
T.Rex
Los huesos de la cadera sirven
como punto de pivote para los huesos de las piernas y ayuda a proporcionar
estabilidad.
Los largos y poderosos miembros
traseros, estaban entre los más largos en proporción con el tamaño corporal de
cualquier terópodo, para compensar el inmenso tamaño del animal, muchos huesos
de su esqueleto eran huecos, reduciendo su peso sin pérdida significativa de
fuerza.
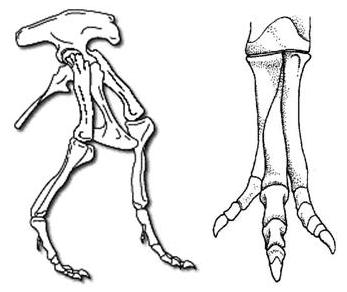
El fémur, la tibia (hueso de la
espinilla) y el peroné (hueso de la pierna) conforman los miembros traceros,
los pies estaban compuestas de muchos huesos, tenían garras (uñas) que eran muy
afilado y puntiagudo. En contraste, los pies de un sauropodos tenían grandes
almohadillas en la parte inferior.
La colas estaba formada por más
de cuarenta vértebras, el propósito de las cola se cree generalmente que para
mantener el equilibrio, proporcionando estabilidad.

El Tiranosaurio es uno de los
pocos dinosaurios cuyo cerebro ha sido estudiado de forma precisa. Es sabido
que este terópodo poseía un cerebro del mismo tamaño que el de un gorila en el
que, al igual que muchas aves de presa, la parte frontal estaba ocupada por
grandes lóbulos olfativos. Esto indica que este dinosaurio podía olfatear una
comida potencial a una distancia de 4 kilómetros o incluso, más. El
Tiranosaurio tenía, además, un gran sentido de la audición que no sólo le
permitía escuchar a sus presas a gran distancia, sino que también le ayudaba a
identificar la dirección de donde provenía un sonido. Estudios recientes
demuestran que el oído interno del Tiranosaurio muestra la capacidad de
detectar ondas de sonido de baja frecuencia y por si fuera poco, estaba
conectado, mediante una serie de nervios, a un tejido almohadillado en la base
de sus patas traseras, lo que le permitía incluso sentir vibraciones leves a
gran distancia. En cuanto a su sentido de la visión, es de notar que sus ojos
estaban posicionados de tal forma que lo dotaban de una excelente visión
binocular. Ésta le proporcionaba una mejor percepción de la distancia entre él
y un punto fijo que aquellos animales cuyos ojos están posicionados en
dirección opuesta. Sin embargo, algunos estudios han demostrado que su nervio
óptico era pequeño con relación al tamaño del cráneo, lo que le dificultaría
cazar en la oscuridad de la noche. Es probable que en estas circunstancias su
visión dependiera principalmente del movimiento, además de valerse de sus demás
sentidos. Respecto a su inteligencia, los restos descubiertos han revelado que
el cerebro de esta criatura era muy pequeño con relación a su enorme cuerpo.
Aún así, muchos científicos deducen que el Tiranosaurio era un animal astuto
que se las ingeniaba para emboscar a sus presas, ya que la astucia y el
intelecto no son factores dependientes del tamaño del cerebro del animal.

Modelo del cerebro de
T.rex
Comportamiento
Se han descubierto fósiles de
T-rex con marcas de mordida causadas por otros de su especie. Algunos de estos
ejemplares muestran signos de sanación, lo que sugiere que sobrevivieron al
ataque, aunque también se han encontrado fósiles con heridas severas que, según
algunos expertos, pudieron haber sido la causa de su muerte. Esto sugiere que
el Tiranosaurio atacaba incluso a otros miembros de su especie y que,
posiblemente, fue un animal territorial.
Sin embargo, hallazgos recientes
confirman que este dinosaurio cuidaba de sus crías. En el año 2000, se
encontraron en Montana fósiles de Tiranosaurio pertenecientes a cinco
individuos de diversa edad, los cuales parecen haber muerto en el mismo lugar,
al mismo tiempo. Basándose en esto, algunos paleontólogos sugieren que es
probable que el T.rex, a pesar de haber sido un animal agresivo que defendía su
territorio, en época de reproducción adoptara un comportamiento grupal. Según
estos expertos, en algunos casos, la familia permanecería unida hasta que las
crías tuvieran la edad suficiente para valerse por sí mismos.
Otras teorías dicen que, en
ciertos casos, el macho era expulsado por la hembra luego de un tiempo, ya que
hay ocasiones en que los machos de algunas especies de carnívoros devoran a su
propia progenie, principalmente, en momentos de desesperación como la estación
seca. Se piensa también que ocasionalmente, los adultos iban en busca de
alimento, dejando a las crías vulnerables ante cualquier posible amenaza. No
obstante, nuevas evidencias apuntan a que el T-rex defendía a sus crías durante
cierto período de tiempo y que incluso les enseñaba a conseguir alimento por sí
mismos, al menos, hasta que ellos lograran hacerlo de forma independiente o
inclusive, durante mucho tiempo después. Es probable que una vez los jóvenes
alcanzaban madurez, los miembros de la manada empezarían a sentirse estresados
por situaciones como la disponibilidad de alimento, lo cual producía
enfrentamientos entre ellos y posiblemente, cada individuo intentaría
expulsarse el uno al otro de la zona de caza hasta que todos tomaban caminos
separados e iban en busca de su propio territorio. Es probable que este
comportamiento familiar no se volviera a repetir hasta la siguiente época de
reproducción.
Otra línea teórica sugiere que el
Tiranosaurio era un animal con un sentido social bastante complejo que, generalmente,
cazaba y se movía en manadas. Los expositores de esta teoría sostienen que los
enfrentamientos entre miembros de la misma especie pudieron deberse a
conflictos por el derecho a la reproducción o bien, a confrontaciones con
miembros de otra manada.

Se ha indicado que los
tiranosaurios rex hembra cuidaban a sus crías hasta que éstas se podían valer
por sí mismas
Depredador o Carroñero

Desde el primer descubrimiento la
mayoría de los científicos han supuesto que el Tyrannosaurus era un depredador,
el mayor carnívoro de su ecosistema, debió haber sido el superpredador, cazando
hadrosáuridos y ceratópsidos, ello no excluye que, al igual que los grandes
depredadores modernos, los tiranosaurios limpiasen cadáveres encontrados
fortuitamente o les robasen presas muertas a otros depredadores si se
presentaba la oportunidad, sin embargo, algunos expertos han sugerido que era
principalmente carroñero. El debate de si Tyrannosaurus fue un depredador
dominante o un carroñero es uno de los más largos en la paleontología.
Varios argumentos apoyan las
diferentes hipótesis:
Las patas delanteras de
Tyrannosaurus son cortas en comparación con las de otros depredadores
conocidos, por lo que no tenían la fuerza de agarre suficiente para aferrar a
su presa.
Tyrannosaurus tenía bulbos
olfatorios y nervios olfativos grandes en relación con su tamaño cerebral. Esto
sugiere un sentido del olfato muy desarrollado, con lo que podría detectar
cadáveres a grandes distancias, como hacen los buitres actuales. Los opositores
de la hipótesis del carroñero puro han usado el ejemplo de los buitres en el
sentido contrario, argumentando que la hipótesis del carroñero es inverosímil
porque los carroñeros puros modernos son sólo grandes aves planeadoras, que
utilizan sus aguzados sentidos y la energía eficiente del planeo para cubrir
amplias zonas con un gasto mínimo de energía. Sin embargo, se ha calculado que
un ecosistema tan productivo como el actual Serengeti podría proporcionar
suficiente carroña para un gran terópodo carroñero, siempre y cuando fuesen de
sangre fría. La ausencia de carroñeros terrestres en los ecosistemas modernos
como Serengeti puede deberse a que las aves planeadoras ahora hacen el trabajo
mucho más eficientemente, mientras que los terópodos grandes no se habrían
enfrentado a esa competencia por su nicho ecológico.

Los dientes de Tyrannosaurus
podían machacar huesos, y por lo tanto podía extraer un máximo de comida,
médula ósea, de los restos de un animal, incluyendo las partes menos
nutritivas. Karen Chin y sus colegas han encontrado fragmentos de hueso en
coprolitos que atribuyen a tiranosaurios, pero señalan que los dientes de un
Tyrannosaurus no estaban bien adaptados a masticar huesos de manera sistemática
para extraer la médula, como hacen las hienas.

Huesos fosilizados de
otros dinosaurios cerca de donde estaría el estómago. Múltiples coprolitos
asignados a Tyrannosaurus rex con huesos rotos.
Dado que al menos algunas de sus
presas potenciales corrían rápido, los indicios de que el Tyrannosaurus caminaba
en lugar de correr sugieren que era carroñero.
Por el contrario, estudios recientes sugieren que el Tyrannosaurus,
aunque era más lento que los grandes depredadores terrestres modernos, bien
pudo haber sido lo suficientemente rápido como para cazar ceratopsianos y
hadrosáuridos grandes.
Otros indicios sugieren un
comportamiento de cazador en el Tyrannosaurus. Sus órbitas oculares están
dispuestas de manera que los ojos miran hacia adelante, dándole una visión
binocular ligeramente mejor que la de los halcones modernos. El linaje de los
Tyrannosaurus tenía una historia de constante mejora de la visión binocular. No
es obvio por qué la selección natural habría favorecido esta tendencia a largo
plazo si los Tyrannosaurus hubiesen sido carroñeros puros, que no habrían
necesitado la percepción avanzada de la perspectiva que proporciona la visión
estereoscópica. En los animales modernos la visión binocular se halla
principalmente en los depredadores pero no en exclusiva, ya que lemúridos y
primates, entre otros no depredadores, también la poseen.
Un esqueleto del hadrosáurido
Edmontosaurus annectens presenta en sus vértebras de la cola una lesión
infligida por un Tyrannosaurus y curada luego. El hecho de que el daño sanase
muestra que el Edmontosaurus sobrevivió al ataque de un Tyrannosaurus durante
su vida, es decir, el Tyrannosaurus había intentado depredación activa. Un
hallazgo similar fue realizado en 2007 y fue descrito por David Burnham en 2013
consistente de dos huesos fusionados de la cola de Edmontosaurus que tenían la
punta de un diente de un Tyrannosaurus adulto incrustada en el hueso, con
evidencia de crecimiento de hueso nuevo que se desarrolló alrededor del diente.
Burnham y sus colegas sugirieron que este hadrosáurido sobrevivió al ataque del
depredador y esto constituye una prueba definitiva de que Tyrannosaurus era un
depredador.

También hay evidencia de una
interacción agresiva entre Triceratops y Tyrannosaurus, ya que aparecen marcas
parcialmente curadas de dientes de Tyrannosaurus en el cuerno frontal y el
escamoso, un hueso del volante del cuello, de un Triceratops, el cuerno mordido
estaba roto, con un crecimiento de hueso nuevo en la fractura. No se sabe cuál
fue la naturaleza exacta de la interacción, cualquiera de los dos animales
podría haber sido el agresor. Al examinar al espécimen Sue, el paleontólogo
Peter Larson encontró una fractura sanada en el peroné y las vértebras de la
cola, cicatrices en los huesos de la cara y un diente de otro Tyrannosaurus
incrustado en una vértebra del cuello. De ser cierto, esto constituiría una
fuerte evidencia de comportamiento agresivo entre Tyrannosaurus, pero no se
sabe si fue competencia por el alimento, por parejas sexuales o canibalismo
activo. Sin embargo, la investigación más reciente de estas supuestas heridas
ha demostrado que la mayoría son infecciones en lugar de lesiones o simplemente
daños en los fósiles ocurridos después de la muerte, y las pocas lesiones
reales son demasiado generales como para probar un conflicto entre individuos
de la misma especie. Un estudio de 2009 demostró que los agujeros en los
cráneos de varios especímenes podrían haber sido causados por parásitos como
las Trichomonas que normalmente infectan a las aves.

Los carnívoros modernos raras
veces son estrictos depredadores o carroñeros. El comportamiento depende de la
disponibilidad de la presa, entre otros factores. Si los tiranosauros eran
carroñeros que practicaban cleptoparasitismo (robo de las presas cazadas por
auténticos depredadores) su masa corporal habría sido un factor intimidante
para ahuyentar a los depredadores, los depredadores coetáneos indiscutibles
como los raptores eran mucho más pequeños y veloces, por lo que la presencia de
un carroñero gigante dotado de grandes dientes les habría hecho huir o
retroceder.
En 2010 se publicaron pruebas de
canibalismo en el género Tyrannosaurus. Se analizaron varios ejemplares de
Tyrannosaurus que presentan en los huesos marcas de dentelladas atribuibles a
otros tiranosaurios. Las marcas de dientes se hallan en el húmero, huesos del
pie y metatarsos, y esto se consideró una prueba de comportamiento carroñero
oportunista, y no de heridas causadas en combate entre miembros de una misma
especie. En una pelea, es de suponer que sería difícil para un T.rex inclinarse
tanto como para llegar a morder en los pies a su rival, por lo que lo más
probable es que las marcas de dientes se hicieran en un cadáver. El que las
marcas aparezcan en partes del cuerpo con cantidades de carne relativamente
escasas sugiere que el Tyrannosaurus se estaba alimentando del cadáver de un
congénere cuyas partes más carnosas ya habían sido devoradas.
La identificación de varios
ejemplares juveniles de tiranosaurio ha permitido a los científicos documentar
los cambios ontogénicos en la especie, estimar su esperanza de vida y
determinar con qué rapidez crecían estos animales. El ejemplar más pequeño
conocido, LACM 28471, llamado el terópodo Jordan, se estima que sólo pesaba 30
kg, mientras que el de mayor tamaño, FMNH PR2081 apodado Sue, probablemente
pesase más de 5.400 kg. El análisis histológico de los huesos de LACM 28471
mostró que sólo tenía dos años cuando murió, mientras que Sue tenía 28 años,
una edad que podría ser cercana al máximo de la especie.
La histología también ha
permitido calcular la edad de otros ejemplares. Las curvas de crecimiento
pueden desarrollarse trazando la masa corporal de diferentes especímenes frente
a su edad. La curva de crecimiento para Tyrannosaurus rex tiene forma de S. Los
jóvenes no pasan de 1.800 kg hasta aproximadamente los 14 años de edad, cuando
el tamaño del cuerpo comienza a aumentar dramáticamente. Durante esta fase de
crecimiento rápido, un joven tiranosaurio debía ganar un promedio de 600 kg al
año durante los próximos cuatro años. A los 18 años de edad, la curva se vuelve
casi horizontal, lo que indica un frenazo drástico del crecimiento. Por
ejemplo, sólo 600 kg separan los 28 años de edad de Sue de los 22 años de un
ejemplar canadiense, el RTMP 81.12.1. Otro estudio histológico reciente
realizado por científicos diferentes corrobora estos resultados, encontrándose
que el crecimiento rápido comenzaba a frenarse alrededor de los 16 años de
edad. Este repentino cambio en la tasa de crecimiento podría ser signo de
madurez física, una hipótesis que es apoyada por el descubrimiento de tejido
medular en el fémur de un tiranosaurio de 16 a 20 años de edad, de Montana, MOR
1125, también conocida como B-rex. El tejido medular se encuentra sólo en las
hembras de aves durante la ovulación, lo que indica que B-rex podría ser una
hembra en su edad reproductiva. La edad de B-rex ha sido estimada en unos 18
años. Otros tiranosáuridos presentan curvas de crecimiento similares, aunque
con menores tasas de crecimiento que resultan en tamaños más pequeños en la
edad adulta.
Más de la mitad de los ejemplares
conocidos de tiranosaurio parecen haber muerto menos de seis años después de
alcanzar la madurez sexual, un patrón que se observa también en otros
tiranosáuridos y actualmente en algunos mamíferos y aves grandes de larga vida.
Estas especies se caracterizan por altas tasas de mortalidad infantil, seguidas
de una mortalidad relativamente baja entre los jóvenes. La mortalidad aumenta
de nuevo después de la madurez sexual, en parte debido a las tensiones de la
reproducción. Un estudio sugiere que la escasez de fósiles juveniles de
Tyrannosaurus rex se debe en parte a las bajas tasas de mortalidad juveniles.
Sin embargo, esta escasez también podría deberse a lo incompleto del registro
fósil o también al sesgo de los recolectores hacia ejemplares fósiles más
grandes y espectaculares.


Existen dudas sobre la existencia
de dimorfismo sexual, es decir diferencias físicas externas significativas
entre machos y hembras, en los Tyrannosaurus.
En los años 1990 el aumento del
número de ejemplares descubierto permitió analizar las diferencias entre
individuos y descubrir lo que parecían ser dos tipos de conformaciones
distintas, llamados morfotipos: uno denominado robusto, construido sólidamente,
y el otro denominado grácil. Se creyó que el morfotipo robusto podría ser
propio de las hembras ya que la mayor amplitud de su pelvis podría haber
servido para permitir el paso de los huevos. Además, se consideró que la
morfología "robusta" estaba correlacionada con un cheurón reducido en
la primera vértebra de la cola, algo que en aquella época se pensaba
erróneamente que en el caso de los cocodrilos también facilitaba la salida de
los huevos.
En los últimos años, los
argumentos a favor de dimorfismo sexual se han debilitado. En 2005 se informó
de que las afirmaciones anteriores sobre dimorfismo sexual en la anatomía del
cheurón de los cocodrilos eran erróneas. Por otra parte se encontró un cheurón
de tamaño natural en la primera vértebra de la cola de Sue, un individuo muy
robusto, lo que indica que de todos modos esta característica no puede
utilizarse para diferenciar los dos morfotipos de Tyrannosaurus rex. Como los
ejemplares de esta especie se han encontrado en un amplio espacio geográfico
que va de Saskatchewan, Canadá a Nuevo México, sudoeste de los Estados Unidos,
podría ser que las diferencias morfológicas entre individuos sean debidas a la
variación geográfica en lugar de al dimorfismo sexual. Las diferencias también
podrían estar relacionadas con la edad, siendo los ejemplares robustos los
animales más viejos.
Solo se ha podido determinar de
manera concluyente el género hembra o macho de un único ejemplar de
Tyrannosaurus, el apodado B-rex. Se ha identificado parte del tejido blando
preservado dentro de sus huesos como tejido medular, un tejido especializado
encontrado exclusivamente en las aves modernas, como fuente de calcio para la
producción de la cáscara de huevo durante la ovulación. Puesto que sólo las
hembras ponen huevos, el tejido medular sólo se encuentra naturalmente en las hembras,
aunque los machos son capaces de producir cuando son inyectados con hormonas
reproductivas femeninas como el estrógeno. Esto sugiere fuertemente que B-rex
era una hembra, y que murió durante la ovulación. La investigación reciente ha
demostrado que el tejido medular no se encuentra en cocodrilos, que se cree que
son los parientes vivos más cercanos de los dinosaurios, además de las aves. La
presencia compartida de tejido medular en las aves y los dinosaurios terópodos
es una prueba más de la estrecha relación evolutiva entre los dos.

Posible postura de
apareamiento del T.rex
No existen pruebas directas ni a
favor ni en contra de que los T. rex hayan tenido plumas. Sin embargo, muchos
de sus parientes próximos sí las tenían y los científicos reconocen la
posibilidad de que ellos también las tuvieran. Se han encontrado restos de
pequeños celurosaurianos, el grupo de dinosaurios al que pertenecen los
tiranosaurios, en la Formación Yixian de Liaoning (China), que presentaban
plumas penáceas o un antiguo pelaje de "protoplumas", lo que sugiere
la posibilidad de que los tiranosáuridos también pudieran haber tenido plumas.
El tiranosauroide antiguo Dilong paradoxus, descubierto en la misma formación,
también mostró filamentos de protoplumas en la cola. Sin embargo, impresiones
de la piel de tiranosaurios adultos de Alberta y Mongolia parecen mostrar las
escamas escaladas típicas de otros dinosaurios. Se ha emitido la hipótesis de
que la presencia de plumas o de escamas podría ser función del tamaño del animal
o de su ubicación geográfica. En climas fríos las plumas habrían sido útiles
como aislamiento térmico, pero no en climas cálidos. Del mismo modo, una
cubierta de plumas habría sobrecalentado a los animales más voluminosos, ya que
en los animales de sangre caliente la cantidad de calor generada es función del
volumen del animal mientras que su refrigeración es función de la superficie
exterior, y el cociente superficie / volumen disminuye cuanto mayor es el
tamaño del animal. Es posible que el T. rex presentara plumas o protoplumas en
otras regiones del cuerpo pero, tal como ocurre con el pelo de los elefantes y
rinocerontes modernos, en áreas reducidas. Las protoplumas podrían haberse
perdido durante la evolución de los tiranosáuridos grandes como Tyrannosaurus,
especialmente en climas cálidos del Cretácico. No se han encontrado fósiles de
T. rex con plumas, pero no porque no las tuviera, sino porque el ambiente donde
se formaron sus fósiles no era propicio para su preservación (como un lago) y
en su lugar, eran zonas de ríos y marismas, donde los sedimentos rápidamente
destruyen los tejidos blandos.
Algunos paleontólogos cree que
las crías de Tiranosaurio rex tenían plumas.

Tampoco está claro si los
tiranosaurios eran ectotérmicos, es decir de sangre fría, o endotérmicos, de
sangre caliente.
Hasta los años 1960 se pensaba
que los tiranosaurios, y la mayoría de los dinosaurios, eran ectotérmicos, de
sangre fría, con un metabolismo de reptil. A partir de entonces científicos
como Robert T. Bakker y John Ostrom sostuvieron que los tiranosaurios debieron
haber sido endotérmicos, de sangre caliente, lo que implica un estilo de vida
muy activo. Los paleontólogos siguen tratando de determinar la capacidad de
Tyrannosaurus para regular su temperatura corporal. Las altas tasas de
crecimiento de los jóvenes Tyrannosaurus rex, medidas por análisis histológico,
son comparables a las de mamíferos y aves y apoyan por tanto la hipótesis de un
metabolismo alto.
Las proporciones de isótopos de
oxígeno en los huesos fosilizados se utilizan a veces para determinar la
temperatura a la que se depositan en el hueso, ya que la relación entre ciertos
isótopos se correlaciona con la temperatura. Un estudio de huesos de
tiranosaurio encontró que las proporciones de isótopos indicaban una diferencia
de temperatura de no más de 4 a 5 °C entre las vértebras del tronco y la tibia
de la pierna. Este rango pequeño de temperatura entre el núcleo del cuerpo y
las extremidades fue usado por el paleontólogo Reese Barrick y el geoquímico William
Showers para indicar que el Tyrannosaurus rex mantenía una temperatura
constante del cuerpo, homeotermia y que disfrutaban de un metabolismo
intermedio entre el de los reptiles ectotérmicos y el de los mamíferos
endotérmicos. Otros científicos han señalado, sin embargo, que la proporción de
isótopos de oxígeno en los fósiles de hoy no presenta necesariamente la misma
relación que en el pasado distante, y puede haber sido alterada durante o
después de fosilización, en el proceso llamado diagénesis. Barrick y Showers
han defendido sus conclusiones en trabajos posteriores, encontrando resultados
similares en otro dinosaurio terópodo de un continente diferente y separado
decenas de millones de años, el Giganotosaurus. Los dinosaurios ornitisquios
también mostraron evidencia de homeotermia, mientras que los varanos de la
misma formación no. Aunque el Tyrannosaurus rex muestre indicios de
homeotermia, ello no significa necesariamente que sea endotérmico. La
termorregulación puede también explicarse por gigantotermia, como ocurre en
algunas especies de tortugas marinas actuales.
Clasificación
Tyrannosaurus es el género tipo,
mundialmente aceptado, de la superfamilia Tyrannosauroidea, la familia
Tyrannosauridae y la subfamilia Tyrannosaurinae. La subfamilia Tyrannosaurinae
incluye al Daspletosaurus de América del Norte y al Tarbosaurus de Asia, los
cuales son, ocasionalmente, clasificados dentro del género del Tyrannosaurus.
Los tiranosáuridos fueron considerados durante mucho tiempo como los
descendientes de grandes terápodos anteriores como los megalosáuridos y los
carnosaurios pero actualmente se les encuadra entre los celurosaurios, que son
generalmente más pequeños.
En 1955 el paleontólogo soviético
Evgeny Maleev nombró al Tyrannosaurus bataar como una nueva especie de
Mongolia. En 1965 esta especie fue renombrada Tarbosaurus bataar. A pesar del
cambio de nombre, el Tarbosaurus de Mongolia a veces es clasificado dentro del
género Tyrannosaurus como T. bataar, aunque la mayor parte de los especialistas
de tiranosaurios, como Tom Holtz, ven suficientes diferencias entre esas dos
especies como para asegurar que se trata de géneros separados, mientras que
otros lo consideran la especie asiática de Tyrannosaurus. Una reciente
descripción del cráneo de Tarbosaurus bataar ha mostrado que es más estrecho
que el de Tyrannosaurus rex y que durante la mordida, la distribución de las
tensiones en los huesos del cráneo eran muy distintas, siendo más cercana a la
de Alioramus, otro tiranosáurido asiático. Un reciente análisis cladistico
encontró que Alioramus, y no el Tyrannosaurus, es el taxón hermano de
Tarbosaurus, lo que sugiere que Tarbosaurus y Tyrannosaurus deben permanecer
separados.
Otros fósiles de tiranosáuridos
encontrados en las mismas formaciones que Tyrannosaurus rex han sido
originalmente atribuidos a diferentes taxones, como el Aublysodon y el
Albertosaurus megagracilis, que fue llamado posteriormente Dinotyrannus
megagracilis en 1995. Sin embargo, en la actualidad estos fósiles son
universalmente considerados como ejemplares juveniles de Tyrannosaurus rex. Un
cráneo pequeño pero muy completo encontrado en Montana, de 60 centímetros de
largo, puede ser una excepción. Este cráneo fue clasificado originalmente como
una especie de Gorgosaurus (G. lancensis) por Charles W. Gilmore en 1946, pero
posteriormente le fue asignado a su propio género, Nanotyrannus. Las opiniones
sobre la validez de N. lancensis están divididas. Muchos paleontólogos
consideran que el cráneo pertenece a un ejemplar juvenil de Tyrannosaurus rex.
Existen diferencias menores entre los dos, incluyendo un mayor número de
dientes en N. lancensis, lo que ha llevado a los científicos a recomendar que
ambos géneros se mantengan separados, hasta que nuevos descubrimientos ayuden a
clarificar estas cuestiones.
Especies
Un gran número de especies
inválidas de Tyrannosaurus ha sido reclasificado bien como T. rex La lista es
la siguiente:
T. gigantus
(1990) nomen nudum, especie no válida, ahora Tyrannosaurus rex.
T. imperiosus
(Osborn, 1905) (originalmente Dynamosaurus) especie no válida, ahora
Tyrannosaurus rex.
T. lancensis
(Gilmore, 1946) (originalmente Gorgosaurus) probablemente Tyrannosaurus rex.
T.
megagracilis (Paul, 1988) (originalmente Albertosaurus) probablemente
Tyrannosaurus rex.
T. stanwinstonorum
(Pickering, 1995) nomen nudum especie no válida, ahora Tyrannosaurus rex.
Hallazgos: Conocido a partir de
numerosos esqueletos parciales en distintos grados de desarrollo ontogenético, hay
más de 30 especímenes de Tyrannosaurus rex identificados, algunos de los cuales
son esqueletos casi completos. Se han encontrado restos fósiles aislados como dientes,
vértebras, falanges, asi como tejido conjuntivo y proteínas en por lo menos uno
de estos especímenes. La abundancia de material fósil ha permitido investigar
en detalle muchos aspectos de su biología, incluyendo su ciclo de vida y su
biomecánica. Los hábitos de alimentación, la fisiología y la velocidad
potencial del Tyrannosaurus rex son objeto de controversia.
Los primeros hallazgos del Tyrannosaurus
rex fueron unos dientes que hoy son documentados como Tyrannosaurus rex fueron
encontrados en 1874 por A. Lakes cerca de Golden (Colorado). A principios de
las década de 1890, J. B. Hatcher recolectó elementos postcraneales en el este
de Wyoming. Estos fósiles se consideraron en un principio pertenecientes a una
especie gigante de Ornithomimus (O. grandis) pero ahora se lo considera un
ejemplar de Tyrannosaurus rex. El primer espécimen, una vértebra parcial, fue
descubierto por Edward Drinker Cope en 1892 y se describió como Manospondylus
gigas. Fue atribuida al Tyrannosaurus rex en 1912 por Henry Fairfield Osborn.
Barnum Brown, el conservador auxiliar del Museo Americano de Historia Natural,
encontró el segundo esqueleto de T. rex en Wyoming en 1900. Este espécimen fue
originalmente nombrado Dynamosaurus imperiosus en el mismo documento en el que
fue descrito el Tyrannosaurus rex. Si no hubiera sido por el orden de las
páginas, Dynamosaurus se habría convertido en el nombre oficial. El material
original del Dynamosaurus se encuentra en las colecciones del Museo de Historia
Natural de Londres.
En total, Barnum Brown encontró
cinco esqueletos parciales del T. rex. Brown recolectó su segundo tiranosaurio
en 1902 y 1905 en la Formación de Hell Creek, Montana. Este fue el holotipo que
Henry Fairfield Osborn, presidente del Museo Americano de Historia Natural, usó
para describir al Tyrannosaurus rex en 1905. En 1941 se lo vendió al Museo
Carnegie de Historia Natural en Pittsburgh (Pensilvania). El cuarto hallazgo de
Brown, que fue el más importante, también descubierto en la Formación de Hell
Creek, está expuesto en el Museo Americano de Historia Natural en Nueva York.
Edward Drinker Cope encontró en
1892 dos vértebras parciales de Tyrannosaurus rex, una de las cuales se
encuentra perdida, y llamó a la especie Manospondylus gigas. Osborn reconoció
las similitudes entre M. gigas y T. rex ya en 1917 pero, debido a la pobre
naturaleza de los restos vertebrales de Manospondylus, no pudo sinonimizarlos.
En junio de 2000, un equipo del
Instituto Black Hills localizó la ubicación del M. gigas en Dakota del Sur y
desenterró nuevos huesos de Tyrannosaurus en dicho lugar. Los investigadores
concluyeron que se trataba del mismo individuo y que los restos eran idénticos
a los de Tyrannosaurus rex. De acuerdo a las reglas de Código Internacional de
Nomenclatura Zoológica (ICZN), el sistema que gobierna los nombres científicos
de los animales, Manospondylus gigas debería haber tenido prioridad sobre
Tyrannosaurus rex por haber sido utilizado primero. Sin embargo la cuarta
edición de la ICZN, que empezó a tener efecto el 1 de enero de 2000, estableció
una excepción que permite seguir considerando a Tyrannosaurus rex como el
nombre válido. Si alguien lo desafiase ante la ICZN, cosa que todavía no ha
ocurrido, muy probablemente sería considerado un nomen protectum ("nombre
protegido") y Manospondylus gigas sería considerado nomen oblitum
("nombre olvidado").
Varios otros esqueletos de
Tyrannosaurus rex fueron descubiertos hasta finales de la década de 1980. El
cráneo de Nanotyrannus, frecuentemente considerado un T. rex juvenil, fue
recuperado de Montana en 1942. En 1966, un grupo de trabajadores del Museo
Americano de Historia Natural bajo la dirección de Harley Garbani descubrió un
cráneo completo de T. rex maduro (LACM 23844). Cuando fue mostrado en Los
Ángeles, LACM 23844 se convirtió en el mayor cráneo expuesto de T. rex en todo
el mundo. Otros cráneos y esqueletos parciales fueron descubiertos en Dakota
del Sur y Alberta (Canadá) a principios de la década de 1980.
Hasta 1987, los restos de
Tyrannosaurus rex eran escasos. Sin embargo, en las décadas de 1980-1990 se ha
presenciado el descubrimiento y la descripción de alrededor de una docena de
especímenes adicionales. El primero fue el Tyrannosaurus apodado Stan, en honor
al paleontólogo aficionado Stan Sacrison, que se encontró en la Formación Hell
Creek cerca de Buffalo (Dakota del Sur) en la primavera de 1987. Después de
30.000 horas de excavación y preparación, surgió un esqueleto completo al 65 %
que ahora se expone en el Museo Black Hills de Historia Natural en Hill City
(Dakota del Sur). Este Tyrannosaurus, cuyo nombre de inventario es BHI 3033,
presenta muchas patologías en sus huesos, incluyendo fracturas en costillas y
cuello que luego sanaron y un espectacular agujero en la parte trasera de su
cabeza, del tamaño de un diente de Tyrannosaurus.
Susan Hendrickson, paleontóloga
amateur, descubrió el esqueleto fósil de T. rex más completo (más del 85%) y de
mayor tamaño conocido hasta ahora, en la Formación de Hell Creek cerca de Faith
(Dakota del Sur), el 12 de agosto de 1990. Sobre la propiedad de ese espécimen
de T. rex, ahora llamado Sue en honor a su descubridora, se entabló una
enconada batalla legal. En 1997 esta se resolvió a favor de Maurice Williams,
dueño original del terreno, y la colección fósil se vendió en subasta por $7,6
millones de dólares. Actualmente el esqueleto se ha vuelto a montar y se expone
en el Museo Field de Historia Natural, en Chicago. El estudio de los huesos
fosilizados de Sue muestra que el espécimen alcanzó su tamaño completo a los 19
años de edad y murió 9 años después, viviendo en total 28 años de edad.

Reconstrucción de Sue

Susan Hendrickson
junto a Sue
Se han descubierto otros dos
fósiles de T. rex en la misma cantera en la que se encontró a Sue, un subadulto
y un juvenil; lo cual indica que el T. rex quizás vivía en manadas u otra clase
de grupos. Las primeras especulaciones de que Sue pudo haber muerto por una
mordedura en la parte posterior de la cabeza no han podido ser confirmadas.
Muchos estudios posteriores han mostrado muchas patologías, pero no se han
encontrado marcas de mordeduras. El daño en la parte posterior del cráneo pudo
haber sido causado por aplastamiento post-mortem. Especulaciones indican que
Sue pudo haber muerto de hambre después de contraer una infección parasitaria
por comer carne putrefacta. La parasitosis resultante habría causado
inflamación en la garganta, impidiendo en última instancia que Sue pudiera
ingerir alimento. Esta hipótesis es apoyada por los agujeros finos y lisos en
su cráneo, que son similares a los causados en los pájaros modernos que
contraen el mismo tipo de parásito.
En el verano boreal de 2000, Jack
Horner descubrió 5 especímenes de Tyrannosaurus cerca de la Reserva de Fort
Peck en Montana. Uno de estos esqueletos, apodado C. rex, fue reportado como el
más largo de los Tyrannosaurus jamás encontrado.
En 2001, el 50 % del esqueleto de
un ejemplar juvenil de tiranosaurio fue descubierto en la Formación de Hell
Creek en Montana por un equipo de investigadores del Museo Burpee de Historia
Natural de Rockford (Illinois). Apodado Jane, el hallazgo fue inicialmente
considerado el primer esqueleto conocido del pequeño tiranosáurido nanotirano
pero una investigación posterior reveló que el fósil pertenecía a un ejemplar
juvenil de tiranosaurio. Este espécimen es el ejemplar juvenil más completo y
mejor preservado hallado hasta la fecha. Jane ha sido examinada por Jack
Horner, Peter Larson, Robert Bakker, Gregorio Erikson y varios otros
paleontólogos de renombre debido a la circunstancia única que constituye la
edad del ejemplar en el momento de su muerte. Jane está actualmente en
exposición en el Museo Burpee de Historia Natural en Rockford (Illinois).
En 2005 se anunció la
recuperación de tejido blando de la cavidad medular de un hueso de la pata
fosilizada de un T. rex, que databa aproximadamente de hace 68 millones de
años. El hueso había sido roto intencionadamente aunque con renuencia para ser
enviado y no fue conservado de la manera habitual porque su descubridora estaba
deseando investigar el tejido blando. Designado como el espécimen MOR 1125 del
Museo de las Rocosas, el dinosaurio había sido desenterrado previamente en la Formación
de Hell Creek. Se pudieron reconocer vasos sanguíneos flexibles y bifurcados y
el tejido de la fibrosa pero elástica matriz del hueso. Además, se encontraron
microestructuras parecidas a las células de la sangre dentro de la matriz y los
vasos sanguíneos. Las estructuras son semejantes a las células y vasos
sanguíneos del avestruz actual. Sin embargo, ya que este material parece haber
sido conservado por un proceso desconocido y distinto al de la fosilización
normal, los investigadores tienen cuidado de no afirmar que este se trate de
material original del dinosaurio.
Un equipo científico ha afirmado
que lo que realmente se encontró en el interior del hueso de tiranosaurio no
era tejido original sino una biopelícula pegajosa creado por bacterias que
cubrían los huecos ocupados originalmente por vasos sanguíneos y células. Sin
embargo no hay evidencia de que una biopelícula pueda producir ramificaciones y
tubos huecos como las observadas en este caso.
Si resultase ser el material
original, cualquier proteína sobreviviente podría usarse para estimar
indirectamente algunos de los contenidos del ADN (ácido desoxirribonucleico) de
los dinosaurios involucrados, porque cada proteína se crea típicamente por un
gen específico. La ausencia de hallazgos anteriores puede ser meramente
consecuencia de que los paleontólogos asumían que la conservación del tejido
era imposible, y simplemente no lo observaron. Desde este hallazgo, se han
encontrado otros dos tiranosaurios y un hadrosaurio que presentaban este tipo de
estructuras y tejidos blandos. La investigación sobre algunos de los tejidos
involucrados ha sugerido que las aves están más cerca de los tiranosaurios en
el árbol evolutivo que de otros animales modernos.
En 2006 la Universidad Estatal de
Montana reveló que estaba en posesión del mayor cráneo de tiranosaurio hallado
hasta ahora. Descubierto en los años 1960 y recientemente reconstruido, el
cráneo mide 149,9 centímetros de largo; comparado con el cráneo de Sue (140,7
cm) es un 6,5% mayor.
Debido a que los pies de los
diferentes terópodos dejan todas huellas de tres dedos muy similares Sin
embargo, se han asignado provisionalmente a Tyrannosaurus dos huellas fósiles
aisladas.
La primera fue descubierta en
Philmont Scout Ranch, Nuevo México, en 1983 por el geólogo estadounidense
Charles Pillmore. Originalmente se pensó que pertenecían a un hadrosáurido, sin
embargo el examen de la huella reveló una gran
huella de talón, desconocida en ornitópodos, y los indicios de lo que
pudo haber sido el espolón, como cuarto dígito del pie de un tiranosaurio. La
huella dio lugar a unos nuevos icnogénero e icnoespecie, Tyrannosauripus
pillmorei, publicados en 1994 por Martin Lockley y Adrian Hunt. Estos autores
sugirieron que era muy probable que fuera hecha por un Tyrannosaurus rex, lo
que la convertiría en la primera huella conocida de esta especie. La huella, de
83 centímetros de largo por 71 de ancho, se imprimió en lo que una vez fue el
lecho fangoso de un humedal con vegetación.

Icnitas de T.rex en Philmont Scout Ranch, Nuevo México, descubierta
en 1983
Una segunda huella que pudo haber
sido hecha por un Tyrannosaurus fue descubierta en 2007 por el paleontólogo
británico Phil Manning, en la Formación Hell Creek de Montana y publicada en
2008 por Manning, Ott y Falkingham. Mide 72 centímetros de largo por 76 de
ancho, más corta y algo más ancha que la descrita por Lockley y Hunt. Los
posibles candidatos a la autoría de esta huella son Tyrannosaurus y
Nanotyrannus, los únicos grandes terópodos conocidos en la Formación Hell
Creek, si bien pudiera pertenecer a algún otro dinosaurio carnívoro aún
desconocido.
No hay comentarios:
Publicar un comentario